При расшифровке генома дрозофилы было установлено что. Полный геном одного биологического вида найден в другом. Перспективы изучения геномов растений
Издательство «БИНОМ. Лаборатория знаний» выпускает книгу воспоминаний ученого-генетика Крейга Вентера «Расшифрованная жизнь». Крейг Вентер известен работами по прочтению и расшифровке генома человека. В 1992 году он основал Институт исследований генома (TIGR). В 2010 году Вентер создал первый в мире искусственный организм – синтетическую бактерию Mycoplasma laboratorium. Мы предлагаем вам ознакомиться с одной из глав книги, в которой Крейг Вентер рассказывает о работе 1999–2000 годов по секвенированию генома мухи дрозофилы.
Вперед, и только вперед
Фундаментальные аспекты наследственности оказались, к нашему удивлению, довольно просты, а потому появилась надежда, что, возможно, природа не так уж непознаваема, а ее не раз провозглашаемая самыми разными людьми непостижимость - просто еще одна иллюзия, плод нашего невежества. Это вселяет в нас оптимизм, поскольку, если бы мир был настолько сложным, как уверяют некоторые наши друзья, у биологии не было бы никакого шанса стать точной наукой.
Томас Хант Морган . Физические основы наследственности
Многие спрашивали меня, почему из всех живых существ на нашей планете я выбрал дрозофилу; других интересовало, почему я сразу не перешел к расшифровке генома человека. Дело в том, что нам нужна была основа для будущих экспериментов, мы хотели быть уверенными в правильности нашего метода, прежде чем потратить почти 100 миллионов долларов на секвенирование генома человека.
Маленькая дрозофила сыграла огромную роль в развитии биологии, особенно генетики. Род дрозофилы включает разных мушек - уксусных, винных, яблочных, виноградных, а также фруктовых, - всего около 26 сотен видов. Но стоит произнести слово «дрозофила», и любой ученый сразу подумает об одном определенном виде - Drosophilamelanogaster. Из-за того, что она быстро и легко размножается, эта крошечная мушка служит для биологов-эволюционистов модельным организмом. Они используют ее, чтобы пролить свет на чудо творения - от момента оплодотворения до становления взрослого организма. Благодаря дрозофилам было сделано немало открытий, в том числе обнаружены гомеобокссодержащие гены, регулирующие общее строение всех живых организмов.
Каждый, изучающий генетику, знаком с опытами на дрозофиле, выполненными Томасом Хантом Морганом, отцом американской генетики. В 1910 году он заметил среди обычных красноглазых мушек мутантов мужского пола с белыми глазами. Он скрестил белоглазую мужскую особь с красноглазой женской особью и обнаружил, что их потомство получилось красноглазым: белоглазость оказалась рецессивным признаком, и теперь мы знаем: чтобы у мушек были белые глаза, нужны две копии гена белоглазости, по одному от каждого родителя. Продолжая скрещивать мутантов, Морган обнаружил, что только у мужских особей проявляется признак белых глаз, и сделал вывод, что этот признак связан с половой хромосомой (Y-хромосомой). Морган и его ученики изучали наследуемые признаки у тысяч плодовых мушек. Сегодня эксперименты с дрозофилой ведутся в лабораториях молекулярной биологии всего мира, где это маленькое насекомое изучают более пяти тысяч человек.
Я на собственном опыте понял всю важность дрозофилы, когда использовал библиотеки ее кДНК генов при исследовании адреналиновых рецепторов и обнаружил у мушки их эквивалент - октопаминовые рецепторы. Это открытие указывало на общность эволюционной наследственности нервной системы мушки и человека. Пытаясь разобраться в библиотеках кДНК мозга человека, я путем компьютерного сопоставления генов человека с генами дрозофилы нашел гены со сходными функциями.
Проект секвенирования гена дрозофилы был запущен в 1991 году, когда Джерри Рубин из Калифорнийского университета в Беркли и Аллен Спредлинг из института Карнеги решили, что настало время приняться за эту задачу. В мае 1998 года 25% секвенирования было уже завершено, и я внес предложение, которое, по словам Рубина, было «слишком хорошим, чтобы от него отказаться». Моя идея была довольно рискованной: тысячам исследователей плодовой мушки из разных стран предстояло пристально изучить каждую букву полученного нами кода, сравнивая ее с высококачественными, эталонными данными самого Джерри, а затем сделать заключение о пригодности моего метода.
Исходный план предполагал завершение секвенирования генома мушки в течение шести месяцев - к апрелю 1999 года, чтобы затем начать атаку на геном человека. Мне казалось, это самый эффектный и всем понятный способ продемонстрировать, что наш новый метод работает. А если у нас ничего не получится, полагал я, то лучше в этом быстро убедиться на примере дрозофилы, чем работая над геномом человека. Но, по правде говоря, полная неудача была бы самым впечатляющим провалом в истории биологии. Джерри тоже рисковал своей репутацией, поэтому все в Celera были полны решимости поддержать его. Я попросил Марка Адамса возглавить нашу часть проекта, и так как у Джерри в Беркли тоже была первоклассная команда, наше сотрудничество шло как по маслу.
Прежде всего встал вопрос о чистоте ДНК, которую нам предстояло секвенировать. Как и люди, мушки различаются на генетическом уровне. Если генетических вариаций в популяции более 2%, и мы имеем 50 различающихся индивидуумов в выбранной группе, то расшифровка оказывается весьма сложной. В первую очередь Джерри пришлось провести инбридинг мушек в максимально возможной степени, чтобы предоставить нам однородный вариант ДНК. Но для обеспечения генной чистоты инбридинга было недостаточно: при извлечении ДНК мушки существовала опасность загрязнения генетическим материалом из клеток бактерий, находящихся в пище мушки или в ее кишечнике. Чтобы избежать этих проблем, Джерри предпочитал извлекать ДНК из мушиных эмбрионов. Но и из клеток эмбрионов приходилось сначала выделять ядра с нужной нам ДНК, чтобы не загрязнять ее внеядерной ДНК митохондрий - «силовых установок» клетки. В результате мы получили пробирку с мутноватым раствором чистой дрозофильной ДНК.
Летом 1998 года команда Хэма, имея такую чистую ДНК мушки, приступила к созданию библиотек ее фрагментов. Сам Хэм больше всего любил разрезать ДНК и соединять внахлест полученные фрагменты, понизив чувствительность своего слухового аппарата, чтобы никакие посторонние звуки не отвлекали его от работы. Создание библиотек должно было положить начало масштабному секвенированию, но пока повсюду раздавались одни только звуки дрели, стук молотков и визжание пил. Рядом постоянно мозолила глаза целая армия строителей, а мы продолжали решать важнейшие проблемы - устранение неполадок в работе секвенаторов, роботов и другого оборудования, пытаясь не за годы, а за считанные месяцы создать с нуля настоящую «фабрику» секвенирования.
Первый секвенатор ДНК модели 3700 был доставлен в Celera 8 декабря 1998 года и встречен c большим восторгом и всеобщим вздохом облегчения. Устройство извлекли из деревянного ящика, поместили в комнату без окон в подвале - его временное пристанище, и сразу приступили к пробным испытаниям. Когда он заработал, мы получили очень качественные результаты. Но эти первые экземпляры секвенаторов работали весьма нестабильно, а некоторые были неисправны с самого начала. С работающими тоже постоянно возникали проблемы, порой чуть ли не ежедневно. Например, в программе управления роботом-манипулятором появилась серьезная ошибка - иногда механическая рука робота на большой скорости выдвигалась над устройством и с размаху врезалась в стену. В результате секвенатор останавливался, и для его починки приходилось вызывать ремонтную бригаду. Некоторые секвенаторы выходили из строя из-за блуждающих лазерных лучей. Для защиты от перегрева использовались ленты из фольги и скотча, поскольку при высокой температуре из последовательностей испарялись окрашенные в желтый цвет фрагменты Gs.
Хотя устройства теперь поставлялись регулярно, около 90% из них с самого начала были неисправны. В некоторые дни секвенаторы вообще не работали. Я твердо верил в Майка Ханкапиллера, однако моя вера сильно поколебалась, когда он стал винить в неудачах наших сотрудников, строительную пыль, малейшие колебания температуры, фазы Луны и так далее. Некоторые из нас от стресса даже поседели.
Не подающие признаков жизни 3700-е, ожидающие отправки обратно в ABI, стояли в кафетерии, и, в конце концов, дошло до того, что нам приходилось обедать практически в «морге» секвенаторов. Я был в отчаянии - ведь мне ежедневно нужно было определенное количество работающих устройств, а именно 230! За примерно 70 миллионов долларов компания ABI обещала предоставить нам или 230 абсолютно исправных устройств, работающих без перебоев целый день, или 460, которые работали хотя бы полдня. Кроме того, Майку следовало удвоить количество квалифицированного технического персонала для незамедлительного ремонта секвенаторов после поломки.
Однако какой интерес заниматься всем этим за те же деньги! К тому же у Майка появился еще один клиент - государственный геномный проект, руководители которого уже начали закупать сотни устройств безо всякого тестирования. Будущее Celera зависело от этих секвенаторов, но Майк, по-видимому, не понимал, что и будущее ABI от них зависело. Конфликт был неизбежен, что и проявилось на важном совещании инженеров ABI и моей команды, состоявшемся в Celera.
После того, как мы сообщили об огромном количестве дефектных приборов и о том, как много времени требуется на исправление поломок секвенаторов, Майк снова попытался свалить всю вину на моих сотрудников, но даже его собственные инженеры с ним не согласились. В конце концов вмешался Тони Уайт. «Мне все равно, сколько это стоит и кого нужно прибить за это», - сказал он. Тогда он в первый и последний раз действительно встал на мою сторону. Он приказал Майку как можно скорее обеспечить поставку новых секвенаторов, даже в ущерб другим клиентам и даже если пока неизвестно, во сколько это обойдется.
Тони также распорядился, чтобы Майк нанял еще двадцать специалистов для оперативного ремонта и определения причин всех проблем. На деле это было легче сказать, чем сделать, потому что опытных работников не хватало. Начать с того, что Эрик Ландер переманил двоих из самых квалифицированных инженеров, и по мнению Майка, тут тоже были виноваты мы. Повернувшись к Марку Адамсу, Майк сказал: «Вы должны были нанять их раньше, чем это сделал кто-то другой». После такого заявления я окончательно потерял к нему всякое уважение. Ведь согласно нашему договору, я не мог нанимать сотрудников ABI, в то время как Ландер и другие руководители государственного проекта генома имели на это право, поэтому очень скоро лучшие инженеры ABI начали работать на наших конкурентов. К концу совещания я понял - проблемы остались, но луч надежды на улучшение все-таки забрезжил.
Так и произошло, хотя и не сразу. Наш арсенал секвенаторов увеличился с 230 до 300 устройств, и если 20–25% из них отказывали, мы все-таки имели около 200 работающих секвенаторов и кое-как справлялись с поставленными задачами. Технические сотрудники работали героически и неуклонно увеличивали темп ремонтных работ, сокращая простои. Все это время я думал об одном: то, что мы делаем, - выполнимо. Неудачи возникали по тысяче причин, но провал не входил в мои планы.
Мы всерьез взялись за секвенирование генома дрозофилы 8 апреля, примерно тогда, когда уже должны были завершить эту работу. Я, конечно, понимал, что Уайт хочет от меня избавиться, но делал все от меня зависящее ради выполнения главной задачи. Напряжение и беспокойство преследовали меня и дома, но с самым своим «доверенным лицом» я эти проблемы обсуждать не мог. Клэр откровенно демонстрировала свое презрение, видя, насколько я поглощен делами Celera. Ей казалось, что я повторяю те же ошибки, которые делал, работая в TIGR/HGS. К 1 июля я чувствовал себя глубоко подавленным, как это уже было во Вьетнаме.
Поскольку конвейерный метод пока у нас не работал, нам предстоял тяжелый изнурительный труд - заново «склеивать» фрагменты генома. Чтобы обнаруживать совпадения и не отвлекаться на повторы, Джин Майерс предложил алгоритм на основе ключевого принципа моего варианта метода дробовика: секвенировать оба конца всех полученных клонов. Поскольку Хэм получал клоны трех точно известных размеров, мы знали, что две концевые последовательности находятся на строго определенном расстоянии друг от друга. Как и прежде, этот способ «нахождения пары» даст нам прекрасную возможность снова собрать геном.
Но поскольку каждый конец последовательности секвенировался отдельно, для обеспечения четкой работы этого метода сборки нужно было вести тщательный учет - для абсолютной уверенности, что мы смогли правильно соединить все пары концевых последовательностей: ведь если хотя бы одна из ста попыток приведет к ошибке и не найдется соответствующая пара для последовательности, все пойдет насмарку и метод не сработает. Один из способов избежать этого - использование штрих-кода и датчиков для отслеживания каждого этапа процесса. Но в начале работы у лаборантов не было необходимого программного обеспечения и оборудования для секвенирования, поэтому приходилось делать все вручную. В Celera небольшая команда, менее двадцати человек, каждый день обрабатывала рекордное количество клонов - 200 тысяч. Мы могли предвидеть некоторые ошибки, например неправильное прочтение данных из 384 лунок, а затем использовать компьютер для нахождения явно ошибочной операции и исправить положение. Конечно, еще оставались отдельные недочеты, но это только подтверждало мастерство команды и уверенность, что мы можем устранять ошибки.
Несмотря на все сложности, мы сумели за четыре месяца прочесть 3156 миллионов последовательностей, всего около 1,76 миллиарда нуклеотидных пар, содержащихся между концами 1,51 миллиона клонов ДНК. Теперь настала очередь Джина Майерса, его команды и нашего компьютера - нужно было сложить все участки вместе в хромосомы дрозофилы. Чем длиннее становились участки, тем менее точным оказывалось секвенирование. В случае дрозофилы последовательности насчитывали в среднем 551 нуклеотидную пару, и средняя точность была 99,5%. Если иметь 500-буквенные последовательности, почти любой может определить места совпадений, передвигая одну последовательность вдоль другой до тех пор, пока не обнаружатся совпадения.
Для секвенирования Haemophilus influenzae у нас было 26 тысяч последовательностей. Для сравнения каждой из них со всеми остальными потребовалось бы проделать 26 тысяч сравнений в квадрате, или 676 миллионов. Геном дрозофилы, с его 3,156 миллиона прочтений потребовал бы около 9,9 триллиона сравнений. В случае человека и мыши, где мы произвели 26 миллионов прочтений последовательности, требовалось около 680 триллионов сравнения. Поэтому не вызывает удивления, что большинство ученых весьма скептически относились к возможному успеху этого метода.
Хотя Майерc и обещал все наладить, у него постоянно возникали сомнения. Теперь он работал дни и ночи напролет, выглядел измученным и как-то посерел. К тому же у него были проблемы в семье, и он стал большую часть свободного времени проводить с журналистом Джеймсом Шривом, который писал о нашем проекте и как тень следил за ходом исследований. Пытаясь как-то отвлечь Джина, я взял его с собой на Карибы - расслабиться и походить под парусом на моей яхте. Но и там он часами сидел, скрючившись над ноутбуком, нахмурив черные брови и щуря свои черные глаза от яркого солнца. И, несмотря на невероятные трудности, Джин и его команда сумели за полгода сгенерировать более полумиллиона строк компьютерного кода для нового ассемблера.
Если бы результаты секвенирования были стопроцентно точными, без повторяющихся ДНК, сборка генома была бы относительно несложной задачей. Но в реальности геномы содержат большое количество повторяющихся ДНК разного типа, разной длины и частоты. С короткими повторами, состоящими из менее пяти сотен пар нуклеотидов, справиться относительно легко, с более длинными повторами - сложнее. Для решения этой проблемы мы использовали метод «нахождения пары», то есть секвенировали оба конца каждого клона и получали клоны разной длины для обеспечения максимального количества совпадений.
Алгоритмы, закодированные в полумиллионе строк компьютерного кода команды Джина, предполагали поэтапный сценарий - от самых «безвредных» действий, например простого перекрывания двух последовательностей, до более сложных, например использования обнаруженных пар для слияния островков перекрывшихся последовательностей. Это было похоже на сложение головоломки, когда небольшие островки собранных участков составляются вместе и образуют большие острова, а затем весь процесс повторяется снова. Только вот в нашей головоломке было 27 миллионов фрагментов. И было очень важно, чтобы участки брались из последовательности высокого качества сборки: представьте себе, что будет, если вы собираете пазл, а цвета или изображения его элементов нечеткие и размытые. Для дальнего порядка последовательности генома значительная доля прочтений должна быть в виде совпадающих пар. Учитывая, что результаты все еще отслеживались вручную, мы с облегчением обнаружили, что 70% имевшихся у нас последовательностей именно такие. Специалисты по компьютерному моделированию объяснили, что при меньшем проценте собрать нашего «шалтая-болтая» было бы невозможно.
И теперь мы смогли использовать ассемблер Celera для секвенирования последовательности: на первом этапе результаты корректировались для достижения самой высокой точности; на втором этапе программа Screener удаляла загрязняющие последовательности из ДНК плазмиды или E. coli. Процесс сборки может быть нарушен всего-навсего какими-то 10 парами оснований «чужой» последовательности. На третьем этапе программа Screener проверяла каждый фрагмент на соответствие известным повторяющимся последовательностям в геноме плодовой мушки - данным Джерри Рубина, который их «любезно» нам предоставил. Местоположение повторов с частично перекрывающимися участками записывалось. На четвертом этапе другая программа (Overlapper) обнаруживала перекрывающиеся участки, сравнивая каждый фрагмент со всеми остальными, - колоссальный эксперимент по обработке огромного объема числовых данных. Ежесекундно мы сравнивали 32 миллиона фрагментов с целью обнаружить по крайней мере 40 перекрывающихся пар оснований с менее 6% различий. При обнаружении двух перекрывающихся участков мы объединяли их в больший фрагмент, так называемый «контиг» - набор перекрывающихся фрагментов.
В идеальном случае этого бы вполне хватило для сборки генома. Но нам приходилось бороться со статтерами и повторами в коде ДНК, а это означало, что один фрагмент ДНК может перекрываться с несколькими различными участками, создавая ложные соединения. Чтобы упростить задачу, мы оставляли только однозначно соединенные фрагменты, так называемые «унитиги». Программа, с помощью которой мы выполняли эту операцию (Unitigger), по существу удаляла всю последовательность ДНК, которую мы не могли с уверенностью определить, оставляя лишь эти унитиги. Этот шаг не только дал нам возможность рассмотреть другие варианты сборки фрагментов, но и существенно упростил задачу. После редукции количество перекрывающихся фрагментов сократилось с 212 миллионов до 3,1 миллиона, и проблема упростилась в 68 раз. Детали головоломки постепенно, но неуклонно вставали на свои места.
А затем мы могли использовать информацию о способе спаривания последовательностей одного и того же клона, используя «каркасный» алгоритм. Все возможные унитиги со взаимно перекрывающимися парами оснований объединялись в специальные каркасы. Для описания этого этапа в своих лекциях я провожу аналогию с детским игрушечным конструктором Tinkertoys. Он состоит из палочек разной длины, которые можно вставлять в отверстия, расположенные на деревянных узловых деталях (шариках и дисках), и составить так объемную конструкцию. В нашем случае узловые детали - это унитиги. Зная, что парные последовательности располагаются на концах клонов длиной в 2 тысячи, 10 тысяч или 50 тысяч пар оснований - то есть как бы находятся на расстоянии определенного количества отверстий друг от друга, - их можно выстроить в одну линию.
В результате тестирования этой методики на последовательности Джерри Рубина, составлявшей примерно одну пятую генома плодовой мушки, мы получили всего лишь 500 пробелов. Проведя в августе испытания на наших собственных данных, мы получили в результате более 800 тысяч небольших фрагментов. Существенно большее количество данных для обработки показало, что методика работала плохо - результат оказался противоположным ожидаемому. В течение нескольких следующих дней паника нарастала, а список возможных ошибок удлинялся. С верхнего этажа корпуса № 2 адреналиновый раж просачивался в комнату, шутливо называемую «Безмятежными покоями». Однако никакого покоя и безмятежности там не ощущалось, особенно в течение по крайней мере пары недель, когда сотрудники буквально кругами слонялись в поисках выхода из создавшегося положения.
В конце концов проблему решил Артур Делчер, работавший с программой Overlapper. Он заметил нечто странное в 678-й строке кода из 150 тысяч строк, в том месте, где пустяковая неточность означала, что важная часть совпадений не записана. Ошибка была исправлена, и 7 сентября у нас было 134 клеточных каркаса, покрывавших действующий (эухроматический) геном плодовой мушки. Мы были в восторге и с облегчением выдохнули. Пришла пора объявить всему миру о нашем успехе.
Конференция по секвенированию генома, которую я начал проводить несколько лет назад, предоставляла для этого прекрасную возможность. Я был уверен, что найдется большое количество жаждущих удостовериться, сдержали ли мы свое обещание. Я решил, что рассказывать о наших достижениях, и прежде всего о процессе секвенирования, сборке генома и значении этого для науки, должны Марк Адамс, Джин Майерс и Джерри Рубин. Из-за наплыва желающих приехать на конференцию мне пришлось перенести ее из Хилтон-Хеда в более вместительный отель «Фонтенбло» в Майами. На конференции присутствовали представители крупных фармацевтических и биотехнических компаний, специалисты по геномным исследованиям со всего мира, довольно много обозревателей, репортеров и представителей инвестиционных компаний - все были в сборе. Наши конкуренты из компании Incyte потратили немалые средства на организацию приема после окончания конференции, корпоративную видеосъемку и прочее - делали все, дабы убедить публику, что именно они предлагают «самую подробную информацию о геноме человека».
Мы собрались в большом конференц-зале. Выдержанный в нейтральных тонах, украшенный настенными светильниками, он был рассчитан на две тысячи человек, но народ все прибывал, и вскоре зал заполнился до отказа. Открытие конференции состоялось 17 сентября 1999 года, и на первом заседании с сообщениями выступили Джерри, Марк и Джин. После небольшого вступления Джерри Рубин объявил, что собравшимся предстоит услышать о лучшем совместном проекте известных компаний, в котором ему когда-либо довелось участвовать. Атмосфера накалялась. Аудитория поняла, что он не стал бы говорить так высокопарно, если бы у нас не было заготовлено что-то действительно сенсационное.
В воцарившейся тишине Марк Адамс начал подробно описывать работу нашего «производственного цеха» в Celera и наши новые методы секвенирования генома. Однако при этом он ни слова не сказал о собранном геноме, словно поддразнивая публику. Затем вышел Джин, поведавший о принципах метода дробовика, о секвенировании Haemophilus, об основных стадиях работы ассемблера. С помощью компьютерной анимации он продемонстрировал весь процесс обратной сборки генома. Отведенное на выступления время заканчивалось, и многие было уже решили, что все ограничится элементарной презентацией с использованием программы PowerPoint, без предъявления конкретных результатов. Но тут Джин c ехидной улыбкой заметил, что аудитория, наверное, захочет все-таки увидеть реальные результаты и не удовольствуется имитацией.
Невозможно было представить наши результаты яснее и выразительнее, чем это сделал Джин Майерс. Он понял, что сами по себе результаты секвенирования не произведут должного впечатления, поэтому для большей убедительности сравнил их с результатами кропотливого исследования Джерри традиционным методом. Они оказались идентичными! Таким образом, Джин сравнил результаты нашей сборки генома со всеми известными маркерами, картированными на геноме плодовой мушки десятки лет назад. Из тысяч маркеров только шесть не совпадали с результатами нашей сборки. Тщательно исследовав все шесть, мы убедились, что секвенирование в Celera было верным и что ошибки содержались в работах, выполненных в других лабораториях старыми методами. Под конец Джин сообщил, что мы только что приступили к секвенированию ДНК человека, и с повторами здесь наверняка будет меньше проблем, чем в случае дрозофилы.
Последовали громкие и продолжительные аплодисменты. Не прекращавшийся и во время перерыва гул означал, что мы своего добились. Кто-то из журналистов заметил участника государственного проекта генома, сокрушенно качающего головой: «Похоже, эти мерзавцы действительно собираются все сделать» 1 . Мы покинули конференцию с новым зарядом энергии.
Оставалось решить две важные проблемы, и обе были нам хорошо знакомы. Первая - как публиковать результаты. Несмотря на подписанный с Джерри Рубином меморандум о взаимопонимании, сотрудники нашего бизнес-отдела не одобряли идею передачи ценных результатов секвенирования дрозофилы в GenBank. Они предлагали разместить результаты секвенирования плодовой мушки в отдельной базе данных в Национальном центре биотехнологической информации, где ими сможет пользоваться каждый при одном условии - не в коммерческих целях. Вспыльчивый, постоянно курящий Майкл Эшбернер из Европейского института биоинформатики был крайне этим недоволен. Он считал, что компания Celera «всех надула» 2 . (Он писал Рубину: «Что, черт подери, происходит в Celera?» 3) Коллинз тоже был недоволен, но что гораздо важнее, недоволен был и Джерри Рубин. В конце концов я все-таки отослал наши результаты в GenBank.
Вторая проблема касалась дрозофилы - у нас были результаты секвенирования ее генома, но мы совершенно не понимали, что они означают. Нужно было проанализировать их, если мы хотели написать статью, - так же, как четыре года назад в случае с Haemophilus. Анализ и описание генома мушки могли занять более года - а у меня такого времени не было, потому что теперь следовало сосредоточиться на геноме человека. Обсудив это с Джерри и Марком, мы решили вовлечь в работу над Drosophila научное сообщество, превратив это в увлекательную научную задачу, и таким образом быстро продвинуть дело, устроить из скучного процесса описания генома веселый праздник - наподобие международного скаутского слета. Мы назвали его «Геномное Джамбори» и пригласили ведущих ученых со всего мира приехать в Роквилл примерно на неделю или дней на десять - проанализировать геном мушки. На основе полученных результатов мы планировали написать серию статей.
Идея всем понравилась. Джерри начал рассылать приглашения на наше мероприятие группам ведущих исследователей, а специалисты по биоинформатике Celera решали, какие компьютеры и программы понадобятся, чтоб сделать работу ученых максимально эффективной. Мы договорились, что Celera оплатит им расходы на проезд и проживание. Среди приглашенных были и самые мои суровые критики, но мы надеялись, что их политические амбиции не повлияют на успех нашей затеи.
В ноябре к нам прибыло около 40 специалистов по дрозофиле, и даже для наших недругов предложение оказалось слишком привлекательным, чтобы от него отказаться. Вначале, когда участники поняли, что им предстоит проанализировать более ста миллионов пар оснований генетического кода в течение нескольких дней, ситуация была довольно напряженной. Пока вновь прибывшие ученые спали, мои сотрудники круглые сутки трудились, разрабатывая программы решения непредвиденных проблем. К концу третьего дня, когда оказалось, что новые программные средства позволяют ученым, как сказал один из наших гостей, «за несколько часов делать потрясающие открытия, на которые раньше уходила чуть ли не вся жизнь», обстановка разрядилась. Ежедневно в середине дня, по сигналу китайского гонга все собирались вместе - обсудить последние результаты, решить текущие проблемы и составить план работы на следующий раунд.
С каждым днем дискуссии становились все увлекательнее. Благодаря Celera, у наших гостей появилась возможность первыми заглянуть в новый мир, и то, что открывалось взору, превосходило ожидания. Скоро оказалось, что нам не хватает времени обсудить все, что хочется, и понять, что все это значит. Марк устроил праздничный ужин, который продолжался очень недолго, так как все быстро устремились обратно в лаборатории. Скоро обеды и ужины поглощались прямо перед экранами компьютеров с выведенными на них данными о геноме дрозофилы. Впервые были обнаружены долгожданные семейства рецепторных генов и одновременно удивительное количество генов плодовой мушки, аналогичных генам болезней человека. Каждое открытие сопровождалось радостными воплями, свистом и дружескими похлопываниями по плечу. Как это ни удивительно, но среди нашего научного пиршества одна пара нашла время для помолвки.
Было, правда, некое опасение: в ходе работы ученые обнаружили всего около 13 тысяч генов вместо ожидаемых 20 тысяч. Поскольку в «непритязательном» черве C. elegans порядка 20 тысяч генов, многие полагали, что у плодовой мушки их должно быть больше, так как у нее в 10 раз больше клеток и даже есть нервная система. Существовал один простой способ удостовериться, что в расчетах нет ошибки: взять 2500 известных генов мушки и посмотреть, сколько их удалось найти в нашей последовательности. После тщательного анализа Майкл Черри из Стэнфордского университета сообщил, что он обнаружил все гены, кроме шести. После обсуждения эти шесть генов были отнесены к артефактам. То, что гены были выявлены без ошибок, воодушевило нас и придало уверенности. Сообщество тысяч ученых, посвятивших себя исследованию дрозофилы, потратили десятки лет, отслеживая эти 2500 генов, а теперь целых 13 600 были перед ними на экране компьютера.
Во время неизбежной фотосессии в конце работы наступил незабываемый момент: после традиционного похлопывания по плечу и дружеских рукопожатий Майк Эшбернер встал на четвереньки, чтобы я увековечил себя на фотографии, поставив ногу на его спине. Так он хотел - несмотря на все свои сомнения и скептицизм - отдать должное нашим достижениям. Известный генетик, исследователь дрозофилы, он даже придумал соответствующую подпись под фотографией: «Стоя на плечах гиганта». (Он отличался довольно тщедушной фигурой.) «Отдадим должное тому, кто этого заслуживает», - написал он позже 4 . Оппоненты наши пытались представить накладки в передаче результатов секвенирования в общедоступную базу данных как отступление от наших обещаний, но и они вынуждены были признать, что слет внес «чрезвычайно ценный вклад в общемировые исследования плодовой мушки» 5 . Испытав, что такое подлинная «научная нирвана», все расстались друзьями.
Мы решили опубликовать три большие статьи: одну по секвенированию всего генома, где Майк будет первым автором, другую - по сборке генома, где первым автором будет Джин, и третью - по сравнительной геномике червя, дрожжей и генома человека с Джерри в качестве первого автора. Статьи были сданы в редакцию Science в феврале 2000 года и опубликованы в специальном выпуске от 24 марта 2000 года, - меньше чем через год после моей беседы с Джерри Рубином в Колд-Спринг-Харборе. 6 Перед публикацией Джерри организовал для меня выступление на ежегодной конференции по исследованиям дрозофилы в Питтсбурге, на которой присутствовали сотни самых видных специалистов в этой области. На каждое кресло в зале мои сотрудники положили компакт-диск, содержащий весь геном дрозофилы, а также оттиски наших статей, опубликованных в Science. Джерри очень тепло представил меня, уверив собравшихся, что я выполнил все взятые на себя обязательства и что мы прекрасно работали вместе. Мое выступление заканчивалось сообщением о некоторых исследованиях, сделанных во время слета, и краткими комментариями к данным на компакт-диске. Аплодисменты после моего выступления вызвали у меня такое же удивление и были так же приятны, как пять лет назад, когда мы с Хэмом впервые представили геном Haemоphilus на съезде микробиологов. Впоследствии статьи по геному дрозофилы стали наиболее часто цитируемыми статьями в истории науки.
Несмотря на то, что тысячи исследователей плодовой мушки всего мира были в восторге от результатов, мои критики быстро перешли в наступление. Джон Салстон назвал попытку секвенирования генома мушки неудачей, хотя полученная нами последовательность была более полной и более точной, чем результат его кропотливой десятилетней работы по секвенированию генома червя, завершение которой потребовало еще четырех лет после публикации чернового варианта в Science. Коллега Салстона Мейнард Олсон назвал последовательность генома дрозофилы «безобразием», в котором «по милости» Celera придется разбираться участникам государственного проекта генома человека. В действительности же команда Джерри Рубина сумела быстро закрыть оставшиеся пробелы в последовательности путем публикации и сравнительного анализа уже расшифрованного генома менее чем через два года. Эти данные подтвердили, что мы допустили 1–2 ошибки на 10 тысяч пар оснований во всем геноме и менее 1 ошибки на 50 тысяч пар оснований работающего (эухроматического) генома.
Однако, несмотря на всеобщее признание проекта Drosophila, летом 1999 года напряженность в наших отношениях с Тони Уайтом достигла апогея. Уайт никак не мог смириться с вниманием, которое пресса уделяла моей персоне. Каждый раз, приезжая в Celera, он проходил мимо развешанных на стенах в коридоре, рядом с моим кабинетом, копий статей о наших достижениях. А тут мы увеличили одну из них - обложку воскресного приложения газеты USA Today. На ней, под заголовком «Удастся ли этому АВАНТЮРИСТУ совершить величайшее научное открытие нашего времени?» 7 был изображен я, в синей клетчатой рубашке, закинув ногу на ногу, а вокруг меня парили в воздухе Коперник, Галилей, Ньютон и Эйнштейн - и никаких признаков Уайта.
Каждый день его пресс-секретарь звонила узнать, нельзя ли Тони принять участие в кажущемся бесконечным потоке интервью, проходящих в Celera. Он немного успокоился - да и то ненадолго, когда на следующий год ей удалось добиться, чтобы его фотографию поместили на обложке журнала Forbes как человека, который смог увеличить капитализацию компании PerkinElmer от 1,5 миллиарда долларов до 24 миллиардов долларов 8 . («Тони Уайт превратил бедолагу PerkinElmer в высокотехнологичного ловца генов».) Тони не давала покоя и моя общественная активность.
Примерно раз в неделю я выступал c докладом, соглашаясь на малую толику из огромного количества приглашений, которые постоянно получал, потому что мир хотел знать о нашей работе. Тони даже жаловался в совет директоров PerkinElmer, переименованную к тому времени в PE Corporation, что мои поездки и выступления нарушают корпоративные правила. Во время двухнедельного отпуска (за свой счет), который я провел в своем доме на Кейп-Код, Тони вместе с финансовым директором Деннисом Уингером и главным юрисконсультом Applera Уильямом Соучем полетел в Celera, чтобы опросить моих ведущих сотрудников насчет «эффективности руководства Вентера». Они надеялись собрать достаточно грязи, чтобы обосновать мое увольнение. Уайт был поражен, когда все сказали, что если я уйду, они тоже уволятся. Это вызвало огромную напряженность в нашей команде, но и одновременно сплотило нас теснее, чем когда-либо. Мы готовы были праздновать каждую победу как последнюю.
После публикации последовательности генома мушки - к тому времени это была самая большая расшифрованная последовательность в истории - Джин, Хэм, Марк и я подняли тост за то, что выдержали Тони Уайта достаточно долго и добились признания наших успехов. Мы доказали, что наш метод будет работать и при секвенировании генома человека. Даже если бы на следующий день Тони Уайт прекратил финансирование, мы знали - наше главное достижение останется с нами. Больше всего на свете я хотел уйти из Celera и не общаться с Тони Уайтом, но поскольку еще больше я хотел секвенировать геном Homo sapiens, мне приходилось идти на компромисс. Я старался, как мог, ублажить Уайта, только бы продолжить работу и завершить задуманное.
Примечания
1. Shreeve J. The Genome War: How Craig Venter Tried to Capture the Code of Life and Save the World (New York: Ballantine, 2005), p. 285.
2. Ashburner M. Won for All: How the Drosophila Genome Was Sequenced (Cold Spring Harbor Laboratory Press, 2006), p. 45.
3. Shreeve J. The Genome War, p. 300.
4. Ashburner M. Won for All, p. 55.
5. Sulston J., Ferry G. The Common Thread (London: Corgi, 2003), p. 232.
6. Adams M. D., Celniker S. E. et al. «The Genome Sequence of Drosophila Melanogaster», Science, № 287, 2185–95, March 24, 2000.
7. Gillis J. «Will this MAVERICK Unlock the Greatest Scientific Discovery of His Age? Copernicus, Newton, Einstein and VENTER?», USA Weekend, January 29–31, 1999.
8. Ross P. E. «Gene Machine», Forbes, February 21, 2000.
Крейг Вентер
Прыгающие гены
В середине прошлого века американская исследовательница Барбара Макклинток обнаружила у кукурузы удивительные гены, способные самостоятельно менять свое положение на хромосомах. Сейчас их называют «прыгающие гены» или транспозабельные (мобильные) элементы. Открытие долгое время не признавали, считая мобильные элементы уникальным явлением, характерным только для кукурузы. Однако именно за это открытие в 1983 году Макклинток была удостоена Нобелевской премии - на сегодня прыгающие гены обнаружены практически у всех изученных видов животных и растений.
Откуда же взялись гены-попрыгунчики, что они делают в клетке, есть ли от них польза? Почему при генетически здоровых родителях семья плодовой мушки дрозофилы из-за прыгающих генов может с большой частотой производить мутантное потомство или даже вовсе оказаться бездетной? Какова роль прыгающих генов в эволюции?
Нужно сказать, что гены, обеспечивающие работу клеток, расположены в хромосомах в определенном порядке. Благодаря этому для многих видов одноклеточных и многоклеточных организмов удалось построить так называемые генетические карты. Однако между генами находится на порядок больше генетического материала, чем в них самих! Какую роль играет эта «балластная» часть ДНК, до конца не установлено, но именно здесь чаще всего и обнаруживают мобильные элементы, которые не только сами перемещаются, но могут прихватывать с собой и соседние фрагменты ДНК.

Откуда ведут свое происхождение гены-попрыгунчики? Предполагают, что по крайней мере часть из них ведет свое происхождение от вирусов, поскольку некоторые мобильные элементы способны формировать вирусные частицы (как, например, мобильный элемент gipsy у плодовой мушки Drosophila melanogaster ). Часть мобильных элементов появляется в геноме путем так называемого горизонтального переноса из других видов. Например, установлено, что мобильный hobo -элемент (в переводе на русский он так и называется - бродяга) Drosophila melanogaster неоднократно заново внедрялся в геном этого вида. Есть версия, что автономность и склонность к «бродяжничеству» могут иметь и некоторые регуляторные участки ДНК.
Полезный балласт
С другой стороны, большая часть прыгающих генов, несмотря на название, ведет себя смирно, хотя и составляет пятую часть от всего генетического материала Drosophila melanogaster или почти половину человеческого генома.
В избыточности ДНК, о которой упоминалось выше, есть свой плюс: балластная ДНК (в том числе и пассивные мобильные элементы) берет на себя удар в случае внедрения в геном чужеродной ДНК. Вероятность того, что новый элемент встроится в полезный ген и тем самым нарушит его работу, снижается, если балластной ДНК гораздо больше, чем значимой.
Некоторая избыточность ДНК полезна так же, как и «избыточность» букв в словах: мы пишем «Мария Ивановна», а говорим «Маривана». Часть букв неизбежно теряется, но смысл остается. Тот же принцип работает и на уровне значимости отдельных аминокислот в молекуле белка-фермента: строго консервативна лишь последовательность аминокислот, формирующая активный центр. Таким образом, на разных уровнях избыточность оказывается своеобразным буфером, обеспечивающим резерв прочности системы. Вот так и мобильные элементы, потерявшие подвижность, оказываются не бесполезными для генома. Как говорится, «с худой овцы хоть шерсти клок», хотя, может быть, здесь лучше бы подошла другая пословица - «каждое лыко в строку».

Мобильные элементы, сохранившие способность прыгать, перемещаются по хромосомам дрозофилы с частотой 10 –2 -10 –5 на ген за поколение в зависимости от типа элемента, генетического фона и внешних условий. Это означает, что один из ста прыгающих генов, находящихся в клетке, после очередного клеточного деления может поменять свою позицию. В результате через несколько поколений распределение мобильных элементов по хромосоме может измениться очень существенно.
Изучать такое распределение удобно на политенных (многонитчатых) хромосомах из слюнных желез личинок дрозофилы. Эти хромосомы во много раз толще обычных, что значительно упрощает их исследование под микроскопом. Как получаются такие хромосомы? В клетках слюнных желез ДНК каждой из хромосом умножается, как при обычном клеточном делении, но сама клетка при этом не делится. В итоге число клеток в железе не меняется, но зато за 10-11 циклов в каждой хромосоме накапливается несколько тысяч одинаковых нитей ДНК.

Отчасти именно благодаря политенным хромосомам прыгающие гены у дрозофилы изучены лучше, чем у других многоклеточных. В результате этих исследований выяснилось, что даже внутри одной популяции дрозофилы трудно найти две особи, которые имеют хромосомы с одинаковым распределением мобильных элементов. Неслучайно считается, что большая часть спонтанных мутаций у дрозофилы вызвана перемещением этих «попрыгунчиков».
Последствия могут быть разными…
По влиянию на геном активные мобильные элементы можно разделить на несколько групп. Часть их выполняет функции, исключительно важные и полезные для генома. Например, теломерная ДНК, расположенная на концах хромосом, у дрозофилы как раз и состоит из особых мобильных элементов. Эта ДНК крайне важна - потеря ее влечет за собой потерю всей хромосомы в процессе клеточного деления, что приводит клетки к гибели.
Другие мобильные элементы - откровенные «вредители». По крайней мере, таковыми их считают на данный момент. Например, мобильные элементы класса R2 могут специфически внедряться в гены членистоногих, кодирующие один из белков рибосом - клеточных «фабрик» по синтезу белка. Особи с подобными нарушениями выживают только потому, что при этом в геноме повреждается лишь часть из множества генов, кодирующих эти белки.

Есть и такие мобильные элементы, которые перемещаются только в репродуктивных тканях, продуцирующих половые клетки. Это объясняется тем, что в разных тканях один и тот же мобильный элемент может производить разные по длине и функции молекулы белка-фермента, необходимого для перемещения.
Примером последних может служить Р-элемент Drosophila melanogaster , попавший в ее природные популяции путем горизонтального переноса из другого вида дрозофил не более ста лет назад. Однако на Земле сейчас вряд ли найдется популяция Drosophila melanogaster , в которой не нашелся бы Р-элемент. При этом надо отметить, что большая часть его копий дефектна, более того - практически везде обнаружен один и тот же вариант дефекта. Роль последнего в геноме своеобразна: он «нетерпим» к своим собратьям и играет роль репрессора, блокируя их перемещение. Так что защита генома дрозофилы от прыжков «чужака» может частично осуществляться его же производными.
Главное - правильно выбрать родителей!
Большая часть прыжков мобильных элементов не сказывается на внешнем виде дрозофилы, потому что приходится на балластную ДНК, но бывают другие ситуации, когда активность их резко возрастает.

Как ни странно, самым мощным фактором, индуцирующим перемещение прыгающих генов, является неудачный подбор родителей. Например, что получится, если скрещивать самок из лабораторной популяции Drosophila melanogaster , которые не имеют Р-элемента (потому что их предки были выловлены из природы около ста лет назад), с самцами, несущими Р-элемент? У гибридов из-за бурного перемещения мобильного элемента может появиться большое количество разнообразных генетических нарушений. Это явление, названное гибридным дисгенезом, вызвано тем, что в материнской цитоплазме отсутствует репрессор, запрещающий перемещение мобильного элемента.
Таким образом, если женихи из популяции А и невесты из популяции Б могут создать многодетные семьи, то обратное не всегда верно. Семья из генетически здоровых родителей может произвести большое количество мутантных или бесплодных потомков, или даже вовсе оказаться бездетной, в случае если папа и мама имеют в геноме разный набор мобильных элементов. Особенно много нарушений появляется, если эксперимент проводить при температуре 29° С. Влияние внешних факторов, накладываясь на генетический фон, усиливает эффект несоответствия геномов, хотя сами по себе эти факторы (даже ионизирующая радиация) в одиночку не способны вызвать столь массовые перемещения мобильных элементов.
Сходные события у Drosophila melanogaster могут произойти с участием и других семейств мобильных элементов.
«Мобильная» эволюция
Клеточный геном можно рассматривать как своего рода экосистему из постоянных и временных членов, где соседи не просто сосуществуют, но и взаимодействуют друг с другом. Взаимодействие хозяйских генов с мобильными элементами пока плохо изучено, но результатов его можно привести множество - от гибели организма в случае повреждения важного гена до восстановления ранее поврежденных функций.
Случается, что и сами прыгающие гены взаимодействуют друг с другом. Так, известно явление, напоминающее иммунитет, когда мобильный элемент не может внедриться в непосредственной близости от уже имеющегося. Однако не все мобильные элементы столь деликатны: например, Р-элементы могут запросто внедряться друг в друга и выводить собратьев из игры.

Кроме того, в геноме существует своего рода саморегуляция числа мобильных элементов. Дело в том, что мобильные элементы могут обмениваться друг с другом гомологичными участками - этот процесс называется рекомбинацией . В результате такого взаимодействия мобильные элементы могут в зависимости от своей ориентации терять (делеция ) или разворачивать (инверсия ) фрагменты хозяйской ДНК, расположенные между ними. Если теряется значительный кусок хромосомы, геном погибнет. В случае инверсии или небольшой делеции создается разнообразие хромосом, что считается необходимым условием для эволюции.
Если рекомбинации происходят между мобильными элементами, расположенными в разных хромосомах, то в результате образуются хромосомные перестройки, которые при последующих клеточных делениях могут привести к несбалансированности генома. А несбалансированный геном, так же как и несбалансированный бюджет, очень плохо делится. Так что гибель неудачных геномов - одна из причин, почему активные мобильные элементы не заполоняют хромосомы безгранично.
Напрашивается естественный вопрос: насколько значим вклад мобильных элементов в эволюцию? Во-первых, большая часть мобильных элементов внедряется, грубо говоря, куда придется, в результате чего они могут повредить или изменить структуру или регуляцию гена, в который внедрились. Тогда естественный отбор отметает неудачные варианты, а удачные варианты с адаптивными свойствами закрепляются.

Если же последствия внедрения мобильного элемента окажутся нейтральными, то такой вариант может сохраниться в популяции, обеспечив некоторое разнообразие структуры гена. Это может пригодиться при неблагоприятных условиях. Теоретически при массовом перемещении мобильных элементов мутации могут появиться во многих генах одновременно, что может оказаться очень полезным при резкой смене условий существования.
Итак, подытожим: мобильных элементов в геноме много и они разные; они могут взаимодействовать как друг с другом, так и с хозяйскими генами; могут вредить и быть незаменимыми. Нестабильность генома, вызванная перемещением мобильных элементов, может закончиться трагедией для особи, но умение быстро меняться - необходимое условие выживания популяции или вида. Благодаря этому создается разнообразие, являющееся базой для естественного отбора и последующих эволюционных преобразований.
Можно провести некоторую аналогию между прыгающими генами и иммигрантами: некоторые иммигранты или их потомки становятся равноправными гражданами, другим дают вид на жительство, третьих - тех, кто не соблюдает законов, - депортируют или сажают в тюрьму. А массовые переселения народов могут быстро изменить само государство.
Литература
Ратнер В. А., Васильева Л. А. Индукция транспозиций мобильных генетических элементов стрессовыми воздейст-виями. Русский переплет. 2000.
Гвоздев В. А. Подвижные ДНК эукариот // Соросовский образовательный журнал. 1998. № 8.
) обнаружили в геноме плодовой мушки (Drosophila ananassae ) полную копию генома бактерии-паразита Wolbachia .
Бактерия вольбахия проживает в цитоплазме клеток хозяина и известна тем, что научилась тонко регулировать размножение, развитие и даже эволюцию своих хозяев. Поэтому её часто называют «микробом-манипулятором» или «повелителем мух» (так как проживает она в клетках насекомых).
Исследование началось с того, что Джули Даннинг-Хотопп (Julie Dunning-Hotopp) из JCVI обнаружила, как некоторые гены вольбахии «кооперируются» с генами дрозофилы, будто они являются частями одного генома.
Майкл Кларк (Michael Clark) – научный сотрудник университета Рочестера — поселил колонию Drosophila ananassae в лаборатории, чтобы вместе с Уэрреном понять, в чём секрет.
Ген вольбахии в геноме дрозофилы (иллюстрация University of Rochester).
«В течение нескольких месяцев, я думал, что в чём-то ошибаюсь, — говорит Кларк, — я даже предположил, что выработалась устойчивость к антибиотику, ведь каждый ген вольбахии я обнаруживал вновь и вновь. Когда же я, наконец, взял ткани, которые оставил в покое несколько месяцев назад, то саму вольбахию не обнаружил».
Сейчас Уэррен и Кларк пытаются понять, в чём преимущество встраивания такого большого куска ДНК для дрозофилы — возможно, «чужие» гены предоставляют хозяину какие-то новые возможности.

А так гены вольбахии переходят в ДНК хозяина (иллюстрация Nicolle Rager Fuller, National Science).
Результаты проведённого исследования опубликованы в статье в журнале Science. В ней авторы предполагают, что горизонтальная передача генов (передача генов между видами, не являющимися родственными) происходит между бактериями и многоклеточными организмами в нашем мире значительно чаще, чем предполагалось ранее.
Расшифровка молекулярно-генетических механизмов манипуляций, осуществляемых вольбахией со своими хозяевами, даст человеку мощные новые средства воздействия на живые организмы и природу в целом.
Впрочем, не все насекомые подвержены плохому влиянию вольбахии. Например, бабочки с островов Самоа "научились" защищать своих самцов. Интересно, научатся ли бороться с нею малярийные комары , которых хотят заразить этой бактерией?
К 50-летию открытия структуры ДНК
А.В. Зеленин
ГЕНОМ РАСТЕНИЙ
А. В. Зеленин
Зеленин Александр Владимирович
- д.б.н.,
заведующий лабораторией Института молекулярной биологии
им. В.А. Энгельгардта РАН.
Впечатляющие достижения программы "Геном человека", а также успехи работ по расшифровке так называемых сверхмалых (вирусы), малых (бактерии, дрожжи) и средних (круглый червь, дрозофила) геномов сделали возможным переход к широкомасштабному изучению крупных и сверхкрупных геномов растений . Насущная необходимость детального изучения геномов наиболее важных в хозяйственном отношении растений была подчеркнута на совещании по геномике растений, состоявшемся в 1997 г. в США [ , ]. За прошедшие с того времени годы достигнуты несомненные успехи в этой области. В 2000 г. появилась публикация о полном секвенировании (установление линейной последовательности нуклеотидов всей ядерной ДНК) генома горчицы малой - арабидопсиса, в 2001 г. - о предварительном (черновом) секвенировании генома риса. Неоднократно сообщалось о работах по секвенированию крупных и сверхкрупных геномов растений (кукуруза, рожь, пшеница), однако эти сообщения не содержали конкретной информации и носили, скорее, характер деклараций о намерении.
Предполагается, что расшифровка геномов растений откроет перед наукой и практикой широкие перспективы. Прежде всего выявление новых генов и цепочки их генетической регуляции позволит существенно повысить продуктивность растений за счет использования биотехнологических подходов. С обнаружением, выделением, размножением (клонированием) и секвенированием генов, отвечающих за такие важнейшие функции растительного организма, как размножение и продуктивность, процессы изменчивости, устойчивости к воздействию неблагоприятных факторов среды, а также гомологичное спаривание хромосом, связывают появление новых возможностей для усовершенствования селекционного процесса. Наконец, выделенные и клонированные гены можно использовать для получения трансгенных растений с принципиально новыми свойствами и анализа механизмов регуляции активности генов.
Важность изучения геномов растений подчеркивает и то обстоятельство, что до настоящего времени число локализованных, клонированных и секвенированных генов растений невелико и колеблется, по различным оценкам, между 800 и 1200. Это в 10-15 раз меньше, чем, например, у человека.
Несомненным лидером в широкомасштабном изучении геномов растений остаются США, хотя интенсивные исследования генома риса проводятся в Японии, а в последние годы и в Китае. В расшифровке генома арабидопсиса, кроме лабораторий США, приняли активное участие исследовательские группы Европы. Явное лидерство США вызывает серьезное беспокойство европейских ученых, которое они ясно выразили на совещании под многозначительным названием "Перспективы геномики в постгеномную эру", состоявшемся в конце 2000 г. во Франции. Опережение американской науки в изучении геномов сельскохозяйственных растений и создании трансгенных растительных форм, по мнению европейских ученых, грозит тем, что в не слишком отдаленном будущем (от двух до пяти десятилетий), когда рост численности населения поставит человечество перед лицом всеобщего продовольственного кризиса, европейская экономика и наука попадут в зависимость от американских технологий. В связи с этим объявлено о создании франко-германской научной программы по исследованию геномов растений ("Plantgene") и вложении в нее значительных средств.
Очевидно, что проблемы геномики растений должны привлечь пристальное внимание российских ученых и организаторов науки, а также руководящих инстанций, поскольку речь идет не только о научном престиже, но и о национальной безопасности страны. Через одно-два десятилетия продовольствие станет важнейшим стратегическим ресурсом.
ТРУДНОСТИ В ИЗУЧЕНИИ ГЕНОМОВ РАСТЕНИЙ
Изучение геномов растений - задача значительно более сложная, чем исследование генома человека и других животных . Это связано со следующими обстоятельствами:
огромными размерами геномов, достигающими для отдельных видов растений десятков и даже сотен миллиардов пар нуклеотидов (п.н.): геномы основных хозяйственно важных растений (кроме риса, льна и хлопка) по размерам либо близки к геному человека, либо превышают его во много раз (таблица);ХРОМОСОМНЫЕ ИССЛЕДОВАНИЯ ГЕНОМОВРезкими колебаниями числа хромосом у различных растений - от двух у некоторых видов до нескольких сотен у других, причем не удается выявить строгой корреляции между размером генома и числом хромосом;
Изобилием полиплоидных (содержащих более двух геномов на клетку) форм с близкими, но не идентичными геномами (аллополиплоидия);
Чрезвычайной обогащенностью геномов растений (до 99%) "незначащей" (некодирующей, то есть не содержащей генов) ДНК, что резко затрудняет стыковку (расположение в правильном порядке) отсеквенированных фрагментов в общий крупноразмерный участок ДНК (контиг);
Неполным (по сравнению с геномами дрозо-филы, человека и мыши) морфологическим, генетическим и физическим картированием хромосом;
Практической невозможностью выделять в чистом виде индивидуальные хромосомы с помощью методов, обычно применяемых с этой целью для хромосом человека и животных (сортировка в потоке и использование гибридов клеток);
Трудностью хромосомного картирования (определение расположения на хромосоме) отдельных генов с помощью гибридизации in situ , обусловленной как высоким содержанием в геномах растений "незначащей" ДНК, так и особенностями структурной организации хромосом растений;
Эволюционной отдаленностью растений от животных, что серьезно осложняет использование для изучения геномов растений сведений, полученных при секвенировании генома человека и других животных;
Длительным процессом размножения большинства растений, что существенно замедляет их генетический анализ.
Хромосомные (цитогенетические) исследования геномов вообще и растений в частности имеют длинную историю. Термин "геном" был предложен для обозначения гаплоидного (единичного) набора хромосом с содержащимися в них генами в первой четверти XX в., то есть задолго до установления роли ДНК как носителя генетической информации .
Описание генома нового, ранее генетически не изученного многоклеточного организма обычно начинают с исследования и описания полного набора его хромосом (кариотипа). Это, разумеется, относится и к растениям, огромное множество которых еще даже не начали изучать.
Уже на заре хромосомных исследований проводили сравнение геномов родственных видов растений на основе анализа мейотической конъюгации (объединения гомологичных хромосом) у межвидовых гибридов. За прошедшие 100 лет возможности хромосомного анализа резко расширились. Сейчас для характеристики геномов растений используют более совершенные технологии: различные варианты так называемого дифференциального окрашивания, позволяющего по морфологическим признакам идентифицировать индивидуальные хромосомы; гибридизацию in situ, дающую возможность локализовать конкретные гены на хромосомах; биохимические исследования клеточных белков (электрофорез и иммунохимия) и, наконец, комплекс методов, основанных на анализе хромосомной ДНК вплоть до ее секвенирования.

Рис. 1. Кариотипы хлебных злаков а - рожь (14 хромосом), б - твердая пшеница (28 хромосом), в - мягкая пшеница (42 хромосомы), г - ячмень (14 хромосом)В течение многих лет изучаются кариотипы хлебных злаков, прежде всего пшеницы и ржи. Интересно, что у разных видов этих растений число хромосом различно, но всегда кратно семи. Отдельные виды хлебных злаков можно надежно распознать по их кариотипу. Например, геном ржи состоит из семи пар крупных хромосом, имеющих на своих концах интенсивно окрашенные гетерохроматические блоки, часто называемые сегментами, или бэндами (рис. 1, а). Геномы пшеницы насчитывают уже 14 и 21 пару хромосом (рис. 1, б, в), причем распределение в них гетерохроматических блоков не такое, как в хромосомах ржи. Различаются между собой и отдельные геномы пшеницы, получившие обозначение А, В и D. Возрастание числа хромосом с 14 до 21 приводит к резкому изменению свойств пшеницы, что нашло отражение в их названиях: твердая, или макаронная, пшеница и мягкая, или хлебная, пшеница. За приобретение мягкой пшеницей высоких хлебопекарных свойств ответственен геном D, содержащий гены белков клейковины, которая придает тесту так называемую всхожесть. Именно этому геному уделяется особое внимание при селекционном улучшении хлебных пшениц. Другой 14-хромосомный злак - ячмень (рис. 1, г) обычно не используют для приготовления хлеба, однако он служит основным сырьем для изготовления таких распространенных продуктов, как пиво и виски.
Интенсивно изучаются хромосомы некоторых дикорастущих растений, используемых для улучшения качества важнейших сельскохозяйственных видов, например диких сородичей пшеницы - эгилопсов . Новые растительные формы создаются путем скрещивания (рис. 2) и отбора. В последние годы значительное усовершенствование методики исследований позволило приступить к изучению геномов растений, особенности кариотипов которых (главным образом мелкие размеры хромосом) делали их ранее недоступными для хромосомного анализа. Так, лишь недавно были впервые идентифицированы все хромосомы хлопка, ромашки и льна .

Рис. 2. Кариотипы пшеницы и гибрида пшеницы с эгилопсом
а - гексаплоидная мягкая пшеница (Triticum astivum ), состоящая из А, В и О геномов; б - тетраплоидная пшеница (Triticum timopheevi ), состоящая из А и G геномов. содержит гены устойчивости к большинству болезней пшеницы; в - гибриды Triticum astivum х Triticum timopheevi , устойчивые к мучнистой росе и ржавчине, отчетливо видно замещение части хромосомПЕРВИЧНАЯ СТРУКТУРА ДНК
По мере развития молекулярной генетики расширилось само понятие генома. Сейчас этот термин трактуется как в классическом хромосомном, так и в осовремененном молекулярном смысле: весь генетический материал отдельного вируса, клетки и организма . Естественно, что вслед за изучением полной первичной структуры геномов (так часто называют полную линейную последовательность оснований нуклеиновых кислот) ряда микроорганизмов и человека на очередь встал вопрос о секвенировании геномов растений.
Из множества растительных организмов для исследования были выбраны два - арабидопсис, представляющий класс двудольных (размер генома 125 млн. п.н.), и рис из класса однодольных (420-470 млн. п.н.). Эти геномы невелики по сравнению с геномами других растений и содержат сравнительно немного повторяющихся участков ДНК. Такие особенности давали надежду на то, что выбранные геномы окажутся доступными для относительно быстрого определения их первичной структуры.

Рис. 3. Арабидопсис - горчица малая - мелкое растение из семейства крестоцветных (Brassicaceae ). На пространстве, равном по площади одной странице нашего журнала, можно вырастить до тысячи индивидуальных организмов арабидопсисаОснованием для выбора арабидопсиса послужили не только небольшие размеры его генома, но и мелкие размеры организма, что позволяет легко выращивать его в лабораторных условиях (рис. 3). Принимали во внимание его короткий репродуктивный цикл, благодаря чему можно быстро проводить опыты по скрещиванию и отбору, детально изученную генетику, легкость осуществления манипуляций со сменой условий произрастания (изменение солевого состава почвы, добавление разных питательных веществ и т.д.) и с испытанием действия на растения различных мутагенных факторов и патогенов (вирусы, бактерии, грибы). Арабидопсис не имеет хозяйственной ценности, поэтому его геном, наряду с геномом мыши, получил название справочного, или, что менее точно, модельного.*
* Появление в отечественной литературе термина "модельный геном" - результат неточного перевода английского словосочетания model genome. Слово "model" означает не только прилагательное "модельный", но и существительное "образец", "эталон", "модель". Правильнее было бы говорить о геноме-образце, или справочном геноме.Интенсивная работа по секвенированию генома арабидопсиса была начата в 1996 г. международным консорциумом, в который вошли научные учреждения и исследовательские группы из США, Японии, Бельгии, Италии, Великобритании и Германии. В декабре 2000 г. стала доступной обширная информация, подводившая итоги определения первичной структуры генома арабидопсиса . Для секвенирования использовали классическую, или иерархическую, технологию: сначала изучали отдельные небольшие участки генома, из которых составляли более крупные участки (контиги), а на финальном этапе - структуру индивидуальных хромосом. Ядерная ДНК генома арабидопсиса распределена между пятью хромосомами. В 1999 г. были опубликованы результаты секвенирования двух хромосом, а появление в печати сведений о первичной структуре остальных трех завершило секвенирование всего генома.
Из 125 млн. пар нуклеотидов определена первичная структура 119 млн., что составляет 92% всего генома. Лишь 8% генома арабидопсиса, содержащих крупные блоки повторяющихся участков ДНК, оказались недоступными для изучения. По полноте и тщательности секвенирования геномов эукариот арабидопсис остается пока в первой тройке чемпионов наряду с одноклеточным дрожжевым организмом Saccharomyces cerevisiae и многоклеточным организмом животного Саеnorhabditis elegance (см. табл.).
В геноме арабидопсиса обнаружено около 15 тыс. индивидуальных генов, кодирующих белки. Приблизительно 12 тыс. из них содержатся в виде двух копий на гаплоидный (единичный) геном, так что общее число генов составляет 27 тыс. Число генов у арабидопсиса не сильно отличается от числа генов у таких организмов, как человек и мышь, однако размеры его генома в 25-30 раз меньше. С этим обстоятельством связаны важные особенности в структуре отдельных генов арабидопсиса и общей структуры его генома.
Гены арабидопсиса компактны, содержат лишь несколько экзонов (участков, кодирующих белки), разделенных короткими (около 250п.н.) некодирующими отрезками ДНК (интронами). Промежутки между отдельными генами составляют в среднем 4.6 тыс. пар нуклеотидов. Для сравнения укажем, что гены человека содержат многие десятки и даже сотни экзонов и интронов, а межгенные участки имеют размеры от 10 тыс. пар нуклеотидов и более. Предполагают, что наличие небольшого компактного генома способствовало эволюционной устойчивости арабидопсиса, поскольку его ДНК в меньшей степени становилась мишенью для воздействия различных повреждающих агентов, в частности, для внедрения в геном вирусоподобных повторяющихся фрагментов ДНК (транспозонов).
Из других молекулярных особенностей генома арабидопсиса следует отметить обогащенность экзонов гуанином и цитозином (44% в экзонах и 32% в интронах) по сравнению с генами животных, а также присутствие дважды повторенных (дуплицированных) генов. Предполагают, что такое удвоение произошло в результате четырех одномоментных событий, заключавшихся в удвоении (повторении) части генов арабидопсиса, или слияния родственных геномов. Эти события, имевшие место 100-200 млн. лет назад, - проявление общей тенденции к полиплоидизации (кратному увеличению числа геномов в организме), характерной для геномов растений. Однако некоторые факты показывают, что у арабидопсиса удвоеннные гены неидентичны и функционируют по-разному, что может быть связано с мутациями в их регуляторных участках.
Еще одним объектом полного секвенирования ДНК стал рис . Геном этого растения тоже невелик (12 хромосом, дающих в сумме 420-470 млн. п.н.), всего в 3.5 раза больше, чем у арабидопсиса. Однако, в отличие от арабидопсиса, рис имеет огромное хозяйственное значение, являясь основой питания для более чем половины человечества, поэтому в улучшении его свойств кровно заинтересованы не только миллиарды потребителей, но и многомиллионная армия людей, активно вовлеченная в весьма трудоемкий процесс его выращивания.
Отдельные исследователи приступили к изучению генома риса еще в 80-х годах прошлого столетия, но серьезного масштаба эти работы достигли лишь в 90-х. В 1991 г. в Японии была создана программа по расшифровке структуры генома риса, объединившая усилия многих исследовательских групп. В 1997 г. на базе этой программы был организован Международный проект "Геном риса" . Его участники решили сконцентрировать усилия на секвенировании одного из подвидов риса (Oriza sativajaponica ), в изучении которого к тому времени уже были достигнуты значительные успехи. Серьезным стимулом и, образно выражаясь, путеводной звездой для такой работы стала программа "Геном человека" .
В рамках этой программы прошла апробацию стратегия "похромосомного" иерархического разделения генома, которую участники международного консорциума использовали при расшифровке генома риса. Однако, если при изучении генома человека с помощью различных приемов выделяли фракции отдельных хромосом, то материал, специфичный для индивидуальных хромосом риса и их отдельных участков, получали методом лазерной микродиссекции (вырезания микроскопических объектов). На предметном стекле микроскопа, где находятся хромосомы риса, под воздействием лазерного луча выжигается все, кроме хромосомы или ее участков, намеченных для анализа. Оставшийся материал используют для клонирования и секвенирования.
Опубликованы многочисленные сообщения о результатах секвенирования отдельных фрагментов генома риса, осуществленного с высокой точностью и детальностью, характерной для иерархической технологии. Считали, что определение полной первичной структуры генома риса будет завершено к концу 2003-середине 2004 г. и результаты вместе с данными по первичной структуре генома арабидопсиса будут широко использоваться в сравнительной геномике других растений.
Однако в начале 2002 г. две исследовательские группы - одна из Китая, другая из Швейцарии и США - опубликовали результаты полного чернового (приблизительного) секвенирования генома риса, выполненного с помощью технологии тотального клонирования . В отличие от поэтапного (иерархического) изучения, тотальный подход основан на одномоментном клонировании всей геномной ДНК в одном из вирусных или бактериальных векторов и получении значительного (огромного для средних и крупных геномов) количества отдельных клонов, содержащих различные отрезки ДНК. На основании анализа этих секвенированных участков и наложения друг на друга идентичных концевых участков ДНК образуется контиг - цепочка стыкованных между собой последовательностей ДНК. Общий (суммарный) контиг представляет собой первичную структуру всего генома или, по крайней мере, индивидуальной хромосомы.
В таком схематичном изложении стратегия тотального клонирования кажется несложной. На деле она встречает серьезные трудности, связанные с необходимостью получения огромного количества клонов (принято считать, что изучаемый геном или его участок должен быть перекрыт клонами, по крайней мере, 10 раз), гигантским объемом секвенирования и чрезвычайно сложной работой по стыковке клонов, требующей участия специалистов по биоинформатике. Серьезным препятствием на пути тотального клонирования служат разнообразные повторяющиеся участки ДНК, число которых, как уже упоминалось, резко возрастает по мере увеличения размера генома. Поэтому стратегию тотального секвенирования используют преимущественно при изучении геномов вирусов и микроорганизмов, хотя она и была успешно применена для исследования генома многоклеточного организма -дрозофилы.
Результаты тотального секвенирования этого генома были "наложены" на огромный массив сведений о его хромосомной, генной и молекулярной структуре, полученных за почти 100-летний период изучения дрозофилы. И все же по степени секвенированности геном дрозофилы (66% общего размера генома) значительно уступает геному арабидопсиса (92%), несмотря на достаточно близкие их размеры - 180 млн. и 125 млн. пар нуклеотидов соответственно. Поэтому недавно предложено называть смешанной технологию, с помощью которой проводилось секвенирование генома дрозофилы.
Для секвенирования генома риса упомянутые выше исследовательские группы взяли два его подвида, наиболее широко культивируемые в азиатских странах, - Oriza saliva L. ssp indicaj и Oriza saliva L. sspjaponica. Результаты их исследований во многом совпадают, но во многом и различаются. Так, представители обеих групп заявили, что ими достигнуто перекрывание контигами приблизительно 92-93% генома. Показано, что около 42% генома риса представлено короткими повторами ДНК, состоящими из 20 пар нуклеотидов, и большинство подвижных ДНК-элементов (транспозонов) находится в межгенных участках. Однако сведения о размерах генома риса существенно различаются.
Для японского подвида размер генома определен равным 466 млн. пар нуклеотидов, а для индийского - 420 млн. Причина такого расхождения не ясна. Оно может быть следствием различных методических подходов в определении размеров некодирующей части геномов, то есть не отражать истинного положения дел. Но не исключено, что 15%-ное различие в размере изученных геномов действительно существует.
Второе серьезное расхождение выявилось в числе обнаруженных генов: для японского подвида - от 46022 до 55615 генов на геном, а для индийского - от 32000 до 50000. Причина такого расхождения не ясна.
Неполнота и противоречивость полученных сведений отмечена в комментариях к опубликованным статьям . Здесь же высказана надежда, что пробелы в знаниях генома риса будут устранены при сопоставлении данных "чернового секвенирования" с результатами детального, иерархического секвенирования, проводимого участниками Международного проекта "Геном риса".
СРАВНИТЕЛЬНАЯ И ФУНКЦИОНАЛЬНАЯ ГЕНОМИКА РАСТЕНИЙ
Полученные обширные данные, половина из которых (результаты китайской группы) общедоступны, несомненно, открывают широкие перспективы как для изучения генома риса, так и для геномики растений в целом. Сравнение свойств геномов арабидопсиса и риса показало, что большая часть генов (до 80%), выявленных в геноме арабидопсиса, обнаружена и в геноме риса, однако приблизительно для половины генов, обнаруженных у риса, пока не удалось найти аналогов (ортологов) в геноме арабидопсиса. В то же время 98% генов, первичная структура которых установлена для других злаков, выявлены в геноме риса.
Вызывает недоумение существенное (почти в два раза) расхождение в числе генов у риса и арабидопсиса. При этом данные черновой расшифровки генома риса, полученные с помощью тотального секвенирования, практически не сопоставлены с обширными результатами изучения генома риса методом иерархического клонирования и секвенирования, то есть не осуществлено то, что сделано в отношении генома дрозофилы. Поэтому остается неясным, отражает ли различие числа генов у арабидопсиса и риса истинное положение дел или же оно объясняется различием в методических подходах.
В отличие от генома арабидопсиса, сведения о генах-двойниках в геноме риса не приведены. Не исключено, что их относительное количество может быть больше у риса, чем у арабидопсиса. В пользу такой возможности косвенно свидетельствуют данные о наличии полиплоидных форм риса. Большей ясности в этом вопросе можно ожидать после завершения Международного проекта "Геном риса" и получения детальной картины первичной структуры ДНК этого генома. Серьезные основания для такой надежды дает тот факт, что после выхода в свет работ о черновом секвенировании генома риса резко увеличилось число публикаций о структуре этого генома, в частности, появились сведения о детальном секвенировании его 1 и 4 хромосом.
Знание, хотя бы приблизительное, числа генов у растений имеет принципиальное значение для сравнительной геномики растений. Вначале считали, что поскольку по своим фенотипическим признакам все цветковые растения весьма близки друг к другу, так же близки должны быть и их геномы. И если мы изучим геном арабидопсиса, то получим сведения и о большинстве геномов других растений. Косвенным подтверждением такого допущения служат результаты секвенирования генома мыши , который удивительно близок к геному человека (около 30 тыс. генов, из которых различными оказалась лишь 1 тыс.).
Можно предположить, что причина различий геномов арабидопсиса и риса кроется в их принадлежности к разным классам растений - двудольным и однодольным. Чтобы прояснить этот вопрос, крайне желательно знать хотя бы черновую первичную структуру еще какого-нибудь однодольного растения. Наиболее реальным кандидатом может стать кукуруза, геном которой приблизительно равен геному человека, но все-таки значительно меньше геномов других злаков. Продовольственное значение кукурузы общеизвестно.
Огромный материал, полученный в результате секвенирования геномов арабидопсиса и риса, постепенно становится основой для широкомасштабного изучения геномов растений методами сравнительной геномики. Такие исследования имеют общебиологическое значение, так как позволяют установить главные принципы организации генома растений в целом и их отдельных хромосом, выявить общие черты структуры генов и их регуляторных участков, рассмотреть соотношение функционально активной (генной) части хромосомы и различных не кодирующих белки межгенных участков ДНК. Сравнительная генетика приобретает все большее значение и для развития функциональной геномики человека. Именно для проведения сравнительных исследований осуществлено секвенирование геномов рыбы фугу и мыши.
Не менее важно изучение отдельных генов, ответственных за синтез индивидуальных белков, определяющих конкретные функции организма. Именно в обнаружении, выделении, секвенировании и установлении функции отдельных генов состоит практическое, в первую очередь медицинское, значение программы "Геном человека". Это обстоятельство несколько лет назад отметил Дж. Уотсон, подчеркнувший, что программа "Геном человека" будет завершена лишь тогда, когда определят функции всех генов человека.
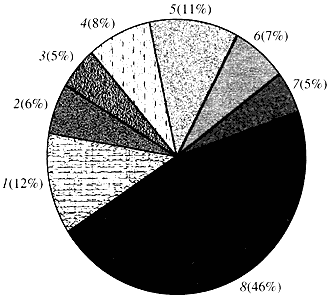
Рис. 4. Классификация по функции генов арабидопсиса
1 - гены роста, деления и синтеза ДНК; 2 - гены синтеза РНК (транскрипция); 3 - гены синтеза и модификации белков; 4 - гены развития, старения и смерти клеток; 5 - гены клеточного метаболизма и энергетического обмена; 6 - гены межклеточного взаимодействия и передачи сигнала; 7 - гены обеспечения прочих клеточных процессов; 8 - гены с неизвестной функциейЧто касается функции генов растений, то мы знаем о них менее одной десятой того, что нам известно о генах человека. Даже у арабидопсиса, геном которого по степени изученности намного превосходит геном человека, функция почти половины его генов остается неизвестной (рис. 4). Между тем у растений, кроме генов, общих с животными, имеется значительное число генов, специфичных только (или, по крайней мере, преимущественно) для них. Речь идет о генах, вовлеченных в транспорт воды и синтез клеточной стенки, отсутствующей у животных, о генах, обеспечивающих образование и функционирование хлоропластов, фотосинтез, фиксацию азота и синтез многочисленных ароматических продуктов. Этот перечень можно продолжить, но уже сейчас ясно, сколь сложная задача стоит перед функциональной геномикой растений.
Полное секвенирование генома дает близкие к истинным сведения об общем количестве генов данного организма, позволяет поместить в банки данных более или менее подробные и достоверные сведения об их структуре, облегчает работу по выделению и изучению индивидуальных генов. Однако секвенирование генома отнюдь не означает установления функции всех генов.
Один из наиболее перспективных подходов функциональной геномики базируется на выявлении работающих генов, на которых идет транскрипция (считывание) мРНК. Этот подход, в том числе использующий современную технологию микрочипов, позволяет одновременно выявлять до десятков тысяч функционирующих генов. Недавно с помощью такого подхода начато изучение геномов растений. Для арабидопсиса удалось получить около 26 тыс. индивидуальных транскриптов, что резко облегчает возможность определения функции практически всех его генов . У картофеля удалось выявить около 20000 тыс. работающих генов, важных для понимания как процессов роста и формирования клубня, так и процессов заболевания картофеля. Предполагается, что это знание позволит повысить устойчивость одного из важнейших пищевых продуктов к возбудителям заболеваний.
Логичным развитием функциональной геномики стала протеомика. Эта новая область науки изучает протеом, под которым обычно подразумевают полный набор белков в клетке в конкретный момент. Такой набор белков, отражающий функциональное состояние генома, все время меняется, тогда как геном остается неизменным.
Изучение белков уже давно используют для суждений об активности геномов растений. Как известно, ферменты, имеющиеся у всех растений, различаются у отдельных видов и сортов последовательностью аминокислот. Такие ферменты, с одинаковой функцией, но разной последовательностью отдельных аминокислот, называют изоферментами. У них разные физико-химические и иммунологические свойства (молекулярная масса, заряд), что можно выявить с помощью хро-матографии или электрофореза. В течение многих лет эти методы успешно использовали для изучения так называемого генетического полиморфизма, то есть различий между организмами, сортами, популяциями, видами, в частности пшеницы и родственных форм злаков. Однако в последнее время в связи с быстрым развитием методов анализа ДНК, включая секвенирование, изучение белкового полиморфизма оказалось замененным исследованием полиморфизма ДНК. Впрочем, прямое изучение спектров запасных белков (проламины, глиадины и др.), определяющих основные питательные свойства злаков, остается важным и надежным способом генетического анализа, селекции и семеноводства сельскохозяйственных растений.
Знание генов, механизмов их экспрессии и регуляции чрезвычайно важно для развития биотехнологии и получения трансгенных растений. Известно, что впечатляющие успехи в этой области вызывают неоднозначную реакцию экологической и медицинской общественности. Однако есть область биотехнологии растений, где эти страхи, если не совсем беспочвенны, то, во всяком случае, представляются малосущественными. Речь идет о создании трансгенных технических растений, не используемых в качестве пищевых продуктов. Недавно в Индии собран первый урожай трансгенного хлопка, устойчивого к ряду заболеваний . Имеются сведения о введении в геном хлопка специальных генов, кодирующих пигментные белки, и получении волокон хлопка, не нуждающихся в искусственном окрашивании. Другая техническая культура, которая может оказаться объектом эффективной генной инженерии, - это лен. Его использование как альтернативы хлопку для получения текстильного сырья обсуждается в последнее время . Эта проблема чрезвычайно важна для нашей страны, утратившей собственные источники хлопкового сырья.
ПЕРСПЕКТИВЫ ИЗУЧЕНИЯ ГЕНОМОВ РАСТЕНИЙ
Очевидно, что структурные исследования геномов растений будут базироваться на подходах и методах сравнительной геномики с использованием в качестве основного материала результатов расшифровки геномов арабидопсиса и риса. Существенную роль в развитии сравнительной геномики растений будут, без сомнения, играть сведения, которые рано или поздно предоставит тотальное (черновое) секвенирование геномов других растений. При этом сравнительная геномика растений будет основываться на установлении генетических взаимосвязей отдельных локусов и хромосом, относящихся к разным геномам. Речь пойдет не столько об общей геномике растений, сколько об избирательной геномике отдельных хромосомных локусов. Так, недавно было показано, что ген, ответственный за яровизацию, расположен в локусе VRn-AI хромосомы 5А гексаплоидной пшеницы и локусе Hd-6 хромосомы 3 риса.
Развитие этих исследований явится мощным толчком к идентификации, выделению и секвени-рованию многих функционально важных генов растений, в частности генов, ответственных за устойчивость к болезням, засухоустойчивость, приспособленность к различным условиям произрастания. Все шире будет использоваться функциональная геномика, основанная на массовом выявлении (скринировании) генов, функционирующих в растениях.
Можно предвидеть дальнейшее совершенствование хромосомных технологий, прежде всего метода микродиссекции. Его использование резко расширяет возможности геномных исследований, не требуя огромных затрат, как, например, тотальное секвенирование геномов. Получит дальнейшее распространение метод локализации на хромосомах растений отдельных генов с помощью гибридизации in situ. В настоящий момент его применение ограничено огромным числом повторяющихся последовательностей в геноме растений, а возможно, и особенностями структурной организации хромосом растений.
Хромосомные технологии в обозримом будущем приобретут большое значение и для эволюционной геномики растений. Эти технологии, относительно недорогие, позволяют быстро оценивать внутри- и межвидовую вариабельность, изучать сложные аллополиплоидные геномы тетраплоидной и гексаплоидной пшеницы, тритикале; анализировать эволюционные процессы на хромосомном уровне; исследовать образование синтетических геномов и введение (интрогрессия) чужеродного генетического материала; выявлять генетические взаимоотношения между индивидуальными хромосомами различных видов.
Изучение кариотипа растений с помощью классических цитогенетических методов, обогащаемых молекулярно-биологическим анализом и компьютерными технологиями, будет использоваться для характеристики генома. Это особенно важно для изучения стабильности и изменчивости кариотипа на уровне не только отдельных организмов, но и популяции, сорта и вида. Наконец, трудно представить, каким образом можно оценить число и спектры хромосомных перестроек (аберрации, мосты) без применения методов дифференциального окрашивания. Такие исследования крайне перспективны для мониторинга окружающей среды по состоянию генома растений.
В современной России вряд ли будет проводиться прямое секвенирование геномов растений. Такие работы, требующие крупных вложений, непосильны для нашей нынешней экономики. Между тем сведений о строении геномов арабидопсиса и риса, полученных мировой наукой и доступных в международных банках данных, достаточно для развития отечественной геномики растений. Можно предвидеть расширение исследований геномов растений, основанных на подходах сравнительной геномики, для решения конкретных задач селекции и растениеводства, а также изучения происхождения различных видов растений, имеющих важное хозяйственное значение.
Можно полагать, что в отечественной селекционной практике и растениеводстве будут широко использоваться такие геномные подходы, как генетическое типирование (RELF, RAPD, AFLP-анализы и т.п.), вполне доступные для нашего бюджета. Параллельно с прямыми методами определения ДНК-полиморфизма для решения проблем генетики и селекции растений будут применяться подходы, основанные на изучении белкового полиморфизма, в первую очередь запасных белков злаков. Широкое применение получат хромосомные технологии. Они относительно недороги, их развитие требует вполне умеренных вложений. В области хромосомных исследований отечественная наука не уступает мировой.
Следует подчеркнуть, что наша наука внесла заметный вклад в становление и развитие геномики растений [ , ].
Основополагающую роль сыграл Н.И. Вавилов (1887-1943).
В молекулярной биологии и геномике растений очевиден пионерский вклад А.Н. Белозерского (1905-1972).
В области хромосомных исследований необходимо отметить работы выдающегося генетика С.Г. Навашина (1857-1930), впервые обнаружившего у растений спутниковые хромосомы и доказавшего, что можно различать отдельные хромосомы по особенностям их морфологии.
Другой классик российской науки Г.А. Левицкий (1878-1942) детально описал хромосомы ржи, пшеницы, ячменя, гороха и сахарной свеклы, ввел в науку термин "кариотип" и развил учение о нем.
Современные специалисты, опираясь на достижения мировой науки, могут внести заметный вклад в дальнейшее развитие генетики и геномики растений.
Автор выражает сердечную благодарность академику Ю.П. Алтухову за критическое обсуждение статьи и ценные советы.Работа коллектива, возглавляемого автором статьи, выполнена при поддержке Российского фонда фундаментальных исследований (гранты № 99-04-48832; 00-04-49036; 00-04-81086), Программы Президента Российской Федерации по поддержке научных школ (гранты № 00-115-97833 и НШ-1794.2003.4) и Программы Российской академии наук "Молекулярно-генетические и хромосомные маркеры в разработке современных методов селекции и семеноводства".
ЛИТЕРАТУРА
1. Зеленин А.В., Бадаева Е.Д., Муравенко О.В.
Введение
в геномику растений // Молекулярная биология. 2001. Т. 35. С. 339-348. 2. Pen E.
Bonanza for Plant Genomics // Science.
1998. V. 282. P. 652-654. 3. Plant genomics // Proc. Natl. Acad. Sci. USA. 1998.
V. 95. P. 1962-2032. 4. Картель Н.А. и др.
Генетика. Энциклопедический
словарь. Минск: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S.
1996. Genome
differentiation in Aegilops. 1. Distribution of highly repetitive DNA sequences
on chromosomes of diploid species // Genome. 1996. V. 39. P. 293-306. История
хромосомного анализа // Биол. мембраны. 2001. Т. 18. С. 164-172.
Образец Всероссийской Проверочной работы по биологии
11 класс
Инструкция по выполнению работы
Проверочная работа включает в себя 14 заданий. На выполнение работы по биологии отводится 1 час 30 минут (90 минут).
Ответами к заданиями являются последовательность цифр, число, слово (словосочетание) или короткий свободный ответ, который записывается в отведенном для этого месте работы. В случае записи неверного ответа зачеркните его и запишите рядом новый.
При выполнении заданий Вы можете пользоваться черновиком. Записи в черновике не учитываются при оценивании работы. Советуем выполнять задания в том порядке, в котором они даны. Для экономии времени пропускайте задание, которое не удается выполнить сразу, и переходите к следующему. Если после выполнения всей работы у Вас останется время, Вы сможете вернуться к пропущенным заданиям.
Баллы, полученные Вами за выполненные задания, суммируются.
Постарайтесь выполнить как можно больше заданий и набрать наибольшее количество баллов.
Пояснения к образцу всероссийской проверочной работы
Приознакомлении с образцом проверочной работы следует иметь в виду, что задания, включенные в образец, не отражают всех умений и вопросов содержания, которые будут проверяться в рамках всероссийской проверочной работы. Полный перечень элементов содержания и умений, которые могут проверяться в работе, приведен в кодификаторе элементов содержания и требований к уровню подготовки выпускников для разработки ВПР по биологии. Назначение образца проверочной работы заключается в том, чтобы дать представление о структуре ВПР, количестве и форме заданий, об уровне их сложности.
1. В опыте экспериментатор осветил часть капли с находящимися в ней амебами. Через непродолжительное время простейшие стали активно двигаться в одном направлении.
1.1. Какое свойство организмов иллюстрирует опыт?
Объяснение: выделяют 7 свойств живых организмов (именно по этим признакам живое отличается от неживого): питание, дыхание, раздражимость, подвижность, выделение, размножение, рост. Амебы из светлой части капли двигаются в темную, так как реагируют на свет, то есть выбираем свойство - раздражимость.
Ответ: раздражимость.
1.2. Приведите пример подобного явления у растений.
Объяснение: здесь можем написать любой пример реакции (проявления раздражимости) у растений.
Ответ: закрытие ловчего аппарата у хищных растений ИЛИ поворот листьев к солнцу или движение подсолнечника в течение дня за солнцем ИЛИ изгибы стеблей из0за изменения ландшафта (окружающей среды).
2. На опушке леса живет и взаимодействуют множество растений, животных, грибов и микроорганизмов. Рассмотрите группу, в которую входят гадюка, орел, ежа сборная, живородящая ящрица, кузнечик обыкновенный. Выполните задания.
2.1. Подпишите изображенные на фотографиях и рисунке объекты, входящие в указанную выше группу.

1 - живородящая ящерица
2 - гадюка
3 - ежа сборная
4 - кузнечик обыкновенный
5 - орел
2.2. Распределите данные организмы по их положению в пищевой цепи. В каждую ячейку запишите номер или название одного из объектов группы.
Пищевая цепь: ежа сборная - кузннечик обыкновенный - живородящая ящерица - гадюка - орел.
Объяснение: пищевую цепь начинаем с продуцента (зеленого растения - производителя органических веществ) - ежа сборная, затем, консумент 1-го порядка (консументы потребляют органические вещества и имеют несколько порядков) - кузнечник обыкновенный, живородящая ящерица (консумент 2-го порядка), гадюка (консумент 3-го порядка), орел (консумнт 4-го порядка).
2.3. Как скажется на численности орлов сокращение количества ежи сборной? Ответ обоснуйте.
Ответ: при сокращении численности ежи сборной уменьшается численность всех последующих компонентов и, в конце концов, орлов, то есть численность орлов снижается.
3. Рассмотрите рисунок, на котором представлена схема круговорота углерода в природе. Укажите название вещества, обозначенного вопросительным знаком.

Объяснение: вопросительным знаком обозначен углекислый газ (СО2), так как при сжигании, дыхании и разложении органических веществ образуется СО2, а при фотосинтезе он образуется (а еще растворяется в воде).
Ответ: углекислый газ (СО2).
4. Петр смешал в 25 пробирках равные количества фермента и его субстрата. Пробирки оставлялись на одинаковое время при различных температурах, измерялась скорость реакции. По результатам эксперимента Петр построил график (по оси х отложена температура (в градусах Цельсия), а по оси у - скорость реакции (в усл. ед.).

Опишите зависимость скорости ферментативной реакции от температуры.
Ответ: при повышении температуры до 30С скорость реакции увеличивается, далее, начинает уменьшаться. Оптимум температуры - 38С.
5. Установите последовательность соподчинения элементов биологических систем, начиная с наибольшего.
Пропущенные элементы:
1. Человек
2. Бицепс
3. Мышечная клетка
4. Рука
5. Аминокислота
6. Белок актин
Запишите соответствующую последовательность цифр.
Объяснение: располагает элементы, начиная с наибольшего уровня:
человек - организменный
рука - органный
бицепс - тканевый
мышечная клетка - клеточный
белок актин - молекулярный (белки состоят из аминокислот)
аминокислота - молекулярный
Ответ: 142365.
6. Белки выполняют множество важных функций в организмах человека и животных: обеспечивают организм строительным материалом, являются биологическими катализаторами или регуляторами, обеспечивают движение, некоторые транспортируют кислород. Для того, чтобы организм не испытывал проблем, человеку в сутки необходимо 100-120 г белков.
6.1. Используя данные таблицы, рассчитайте количество белков, которое человек получил во время ужина, если в его рационе было: 20 г хлеба, 50 г сметаны, 15 г сыра и 75 г трески. Ответ округлите до целых.
Объяснение: в 100 г хлеба содржится 7,8 г белков, тогда в 20 г хлеба в 5 раз меньше белков - 1,56 г. В 100 г сметаны содержится 3 г белка, тогда в 50 г в 2 раза меньше - 1,5 г. В 100 г сыра - 20 г белка, в 15 г сыра - 3 г, в 100 г трески - 17,4 г белка, в 75 г трески - 13,05 г.
Итого: 1,56 + 1,5 + 3 + 13,05 = 19, 01 (что примерно равно 19).
Ответ: 19 г.
ИЛИ
6.1.Человек выпил чашку крепкого кофе, содержащую 120 мг кофеина, который полностью всосался и равномерно распределился по крови и другим жидкостям тела. У исследуемого человека объем жидкостей тела можно считать равным 40 л. Рассчитайте, через какое время (в ч) после приема кофеин перестанет действовать на этого человека, если кофеин перестает действовать при концентрации 2 мг/л, а концентрация его снижается за час на 0,23 мг. Ответ округлите до десятых.
Объяснение: 120 мг кофеина распределились по организму человека в объеме 40 л, то есть концентрация стала 3 мг/л. При концентрации 2 мг/л кофеин перестает действовать, то есть действует только 1 мг/л. Чтобы узнать количество часов, разделим 1 мг/л на 0,23 мг (снижение концентрации в час), получим 4,3 часа.
Ответ: 4,3 часа.
6.2. Назовите один из ферментов, вырабатываемый железами пищеварительной системы:
Ответ: стенки желудка вырабатывают пепсин, который в кислой среде расщепляет белки до дипептидов. Липаза расщепляет липиды (жиры). Нуклеазы расщепляют нуклеиновые кислоты. Амилаза расщепляет крахмал. Мальтаза расщепляет мальтозу до глюкозы. Лактаха расщепляет лактозу до глюкозы и галактозы. Нужно написать один фермент.
7. Определите происхождение болезней, приведенных в списке. Запишите номера каждой из болезней в списке в соответствующую ячейку таблицы. В ячейках таблицы может быть записано несколько номеров.
Список болезней человека:
1. Гемофилия
2. Ветряная оспа
3. Цинга
4. Инфаркт миокарда
5. Холера
Объяснение: см. Болезни человека для ВПР
8. В медицинской генетике широко используется генеалогический метод. Он основан на составлении родословной человека и изучении наследования того или иного признака. В подобных исследованиях используются определенные обозначения. Изучите фрагмент родословного дерева одной семьи, у некоторых членов которой сросшаяся мочка уха.

Используя предложенную схему, определите доминантным или рецессивным является данный признак и сцеплен ли он с половыми хромосомами.
Объяснение: признак является рецессивным, так как в первом поколении не проявляется совсем, а во втором поколении проявляется только у 33% детей. Признак с полом не сцеплен, так как проявляется и у мальчиков и у девочек.
Ответ: рецессивен, с полом не сцеплен.
9. Владимир всегда хотел иметь жёсткие волосы, как у его папы (доминантный признак (А)). Но волосы у него были мягкие, как у мамы. Определите генотипы членов семьи по признаку качества волос. Ответы занесите в таблицу.
Объяснение: мягкие волосы - рецессивный признак (а), отец по данному признаку гетерозиготен, так как сын гомозиготен рецессивен (аа), как и мать. То есть:
Р: Аа х аа
Г: А, а х а
F1: Аа - 50% детей с жесткими волосами
аа - 50% детей с мягкими волосами.
Ответ:
| Мать | Отец | Сын |
| аа | Аа | аа |
10. Екатерина решила сдать кровь в качестве донора. При заборе крови выяснилось, что у Екатерины III группа. Екатерина знает, что у ее матери I группа крови.

10.1. Какой группы может быть кровь у отца Екатерины?
Объяснение: исходя из данных таблицы, у отца Екатерины может быть III или IV группа крови.
Ответ: III или IV.
10.2. Руководствуясь правилами переливания крови, определите, может ли Екатерина быть донором крови для своего отца.

Объяснение: Екатерина с I группой крови является универсальным донором (при условии совпадения резус-факторов), то есть от нее можно перелить кровь отцу.
Ответ: может.
11. Функцией изображенного на рисунке органоида является окисление органических веществ и запасание энергии при синтезе АТФ. В этих процессах важную роль играет внутренняя мембрана этого органоида.

11.1. Как называется этот органоид?
Ответ: на рисунке изображена митохондрия.
11.2. Объясните, как упаковка внутренней мембраны в органоиде связана с выполняемой им функцией.
Отвте: при помощи складок внутренней мембраны увеличивает внутреннюю поверхность органоида и может окислиться большее окличество органических веществ, а также выработаться большее количество АТФ на АТФ-синтазах - ферментативных комплексах, вырабатывающих энергию в виде АТФ (главной энергетической молекулы).
12. Фрагмент иРНК имеет следующую последовательность:
УГЦГААУГУУУГЦУГ
Определите последовательность участка ДНК, послужившего матрицей для синтеза этой молекулы РНК, и последовательность белка, которая кодируется этим фрагментом иРНК. При выполнении задания воспользуйтесь правилом комплементарности и таблицей генетического кода.

Правила пользования таблицей
Первый нуклеотид в триплете берется из левого вертикального ряда; второй - из верхннего горизонтального ряда и третий - из правого вертикального. Там, где пересекутся линии, идущие от всех трех нуклеотидов, и находится искомая аминокислота.
Объяснение: разделим последовательность на триплеты (по три нуклеотида): УГЦ ГАА УГУ УУГ ЦУГ. Запишем соответствующую последовательность нуклеотидов в ДНК (обратную комплементарную последовательность нуклеотидов, учитыва, что А-Т (в РНК У), Г-Ц.
То есть цепь ДНК: АЦГ ЦТТ АЦА ААУ ГАУ.
По последовательности РНК находим соответствующую последовательность аминокислот. Первая аминокислота - цис, далее глу, цис, лей, лей.
Белок: цис-глу-цис-лей-лей.
12.3. При расшифровке генома томата было установлено, что во фрагменте молекулы ДНК доля тимина составляет 20%. Пользуясь правилом Чаргаффа, описывающим количественные соотношения между различными типами азотистых оснований в ДНК (Г+Т = А+Ц), рассчитайте количество (в %) в этой пробе нуклеотидов с цитозином.
Объяснение: если количество тимина - 20%, то количество аденина тоже 20% (так как они комплементарны). На гуанин и цитозин остается 60% (100 - (20 + 20)), то есть по 30%.
Ответ: на цитозин приходится 30%.
13. Современную эволюционную теорию можно представить в виде следующей схемы.

Ответ: вероятно предки жирафа имели разную длину шеи, но так как жирафам нужно было дотягиваться до высоко растущих зеленых листьев, выживали жирафы только с длинной шеей, то есть наиболее приспособленные (данный признак прикреплялся из поколения в поколение, это привело к изменению генетического состава популяции). Таким образом, в ходе естественного отбора выжили только особи с наиболее длинной шеей и длина шеи постепенно увеличивалась.
14. На рисунке изображен кордаит - вымершее древесное голосеменное растение, обитавшее 370-250 млн лет назад.

Используя фрагмент геохронологической таблицы, определите эру и периоды, в которых обитал данный организм. Какие растения были их возможными предками?
Геохронологическая таблица

Объяснение: голосеменные, вероятно, появились в Палеозойскую эру. периоды: Пермь, Карбон (возможно, Девон). Возникли от древовидных папоротников (в палеозойскую эру достигли расцвета более примитивные растения, а голосеменные широко распространились и достигли расцвета в Мезозойскую эру).
Эра: Палеозойская
Периоды: Пермь, Карбон, Девон
Возможные предки: древовидные папоротники
2 018 Федеральная служба по надзору в сфере образования и науки Российской Федерации